






Constituição do Sistema Auditivo Humano
Projeto Ciência Viva
(Ministério da Ciência e da Tecnologia – Portugal)
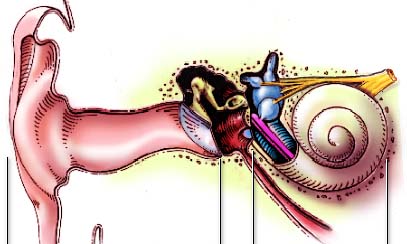
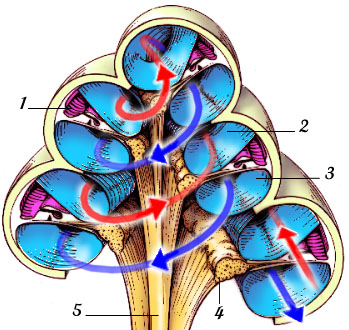
O ouvido humano pode ser separado em três grandes partes, de acordo com a função desempenhada e a localização. São elas: o ouvido externo, o ouvido médio e o ouvido interno. Segue-se então uma vista panorâmica do sistema auditivo humano na qual as suas três zonas constituintes são discriminadas.
|
|
|
autores: Stephan Blatrix, Rémy Pujol |
|
| Figura 1 – Vista panorâmica. |
Fazem parte do ouvido externo o pavilhão auricular e o canal auditivo, cujas funções são recolher e encaminhar as ondas sonoras até ao tímpano. É também no canal auditivo que se dá a produção de cera, que não é mais do que uma forma de este se manter úmido e limpo. Isto porque a cera ajuda a reter partículas de pó, sujidade e microorganismos. Será importante referir que os vulgares cotonetes não devem ser introduzidos no canal auditivo. Isto porque ajudam a empurrar a cera contra o tímpano podendo danificá-lo ou, no mínimo, formar uma barreira que dificulta a audição.
O pavilhão auricular é muito desenvolvido em muitas das espécies de mamíferos terrestres (sendo fundamental na localização de presas e de predadores) e é dotado de movimento. Com a evolução da nossa espécie, essa capacidade foi-se perdendo. Contudo, existem humanos que ainda hoje conseguem produzir pequenos movimentos com as orelhas.
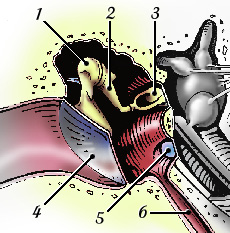
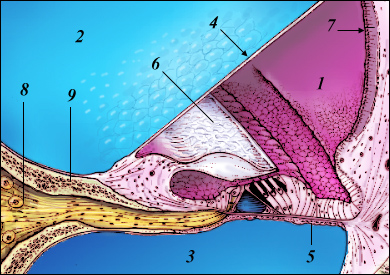
O ouvido médio, também denominado de caixa timpânica, representado com algum detalhe na Figura 2, é uma cavidade com ar, por detrás da membrana do tímpano (4), através da qual a energia das ondas sonoras é transmitida, do ouvido externo até à janela oval na cóclea, esta já no ouvido interno. Essa transmissão de energia é efetuada através de três ossos minúsculos (o martelo (1), a bigorna (2) e o estribo (3)), que vibram, solidários com o tímpano. Estes três ossos (seis, se contarmos com os dois ouvidos) são os mais pequenos que podemos encontrar no corpo humano. No ouvido médio existe ainda um canal, em parte ósseo, em parte fibrocartilagíneo, denominado de trompa de Eustáquio (6), que o mantém em contacto com a rinofaringe. Esta é a forma encontrada pela natureza de manter uma pressão constante no ouvido médio. Para que isso possa acontecer, a trompa de Eustáquio abre e fecha constantemente.
A membrana do tímpano é, na realidade, constituída por três camadas, sendo a camada exterior uma continuação da pele do canal auditivo. A parcela superior da membrana denomina-se de pars flaccida, enquanto que a parcela inferior se chama pars tensa. É na parte central da pars tensa que se localiza a área vibrante ativa, em resposta a um estímulo sonoro. A membrana timpânica é uma estrutura auto-regenerativa, sendo por isso capaz de corrigir um furo na sua estrutura.
A cadeia de pequenos ossos, as suas articulações e ligamentos estão revestidos por uma mucosa e pode tornar-se mais ou menos tensa, pela ação de dois pequenos músculos, o do martelo e o do estribo. Através deste mecanismo é possível limitar a transmissão de energia para o interior da cóclea (algo que é útil para evitar danos no ouvido interno quando estamos expostos a sons de intensidade elevada).
|
|||||||||
| autores: Stephan Blatrix, Rémy Pujol | |||||||||
| Figura 2 – Ouvido médio em mais detalhe. |
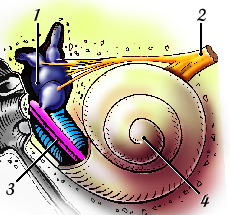
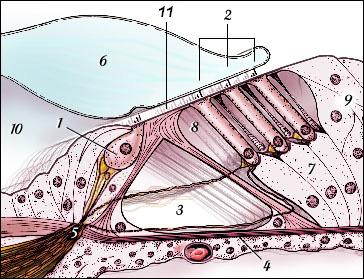
É no ouvido interno ou labirinto que se encontra a parte mais importante do ouvido periférico (o que se encontra entre o pavilhão auricular e os nervos auditivos). É ela a cóclea, em forma de caracol e responsável em grande parte pela nossa capacidade em diferenciar e interpretar sons. De fato, desenrola-se na cóclea uma função complexa de conversão de sinais, em resultado da qual os sons nela recebidos (do tipo mecânico) são transformados em impulsos elétricos que “caminham” até ao cérebro pelo nervo auditivo, onde são depois decodificados e interpretados. Como se pode ver na Figura 3, a cóclea parece uma concha do mar, sendo constituída por um “tubo” ósseo enrolado sobre si próprio, com as dimensões aproximadas de uma ervilha.
|
|||||||
| autores: Stephan Blatrix, Rémy Pujol | |||||||
| Figura 3 – Ouvido interno em mais detalhe. |
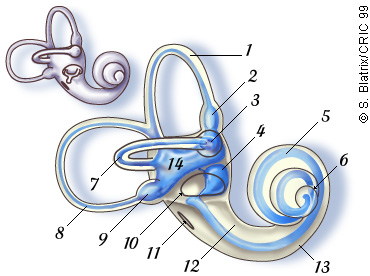
Para além da cóclea, no ouvido interno encontra-se também o labirinto vestibular, constituído pelo sáculo (4) e pelo utrículo (14), que são os órgãos do sentido do equilíbrio e que informam o nosso cérebro sobre a posição do corpo no espaço. Repare-se na figura seguinte. Os canais semicirculares laterais, anteriores, e posteriores fazem também parte do labirinto vestibular, informando o cérebro sobre o movimento rotatório no espaço. A informação proveniente do labirinto vestibular e da cóclea é transmitida ao cérebro pelo nervo auditivo, como ser pode verificar na Figura 3.
|
|||||||||||||||||
| autores: Stephan Blatrix, Rémy Pujol | |||||||||||||||||
| Figura 4 – A cóclea e os canais semicirculares. |
O “tubo” ósseo enrolado que constitui a cóclea encontra-se dividido em toda a sua extensão em três secções, estando todas preenchidas com um fluído semelhante à água. A primeira denomina-se de scala vestibuli (rampa vestibular) e está ligada à janela oval, enquanto que a última, a scala tympani (rampa timpânica) se encontra ligada à janela redonda. Estas duas secções unem-se apenas no fim da cóclea, no chamado helicotrema, e estão separadas por uma terceira secção denominada de ducto coclear. A separação entre as três secções referidas é efetuada por duas membranas. Assim a separar a scala vestibuli do ducto coclear encontra-se a membrana de Reissner e entre o ducto coclear e a scala tympani está a membrana basilar. A membrana basilar é muito importante pois é ela que suporta o órgão de Corti. No órgão de Corti localizam-se as células ciliadas que, quando agitadas pelas vibrações sonoras, produzem impulsos elétricos que o cérebro decodificará. Na figura seguinte podemos observar um corte transversal da espiral coclear, sendo de notar as três secções referidas.
|
||||||||
| autores: Stephan Blatrix, Rémy Pujol | ||||||||
| Figura 5 – A cóclea seccionada. |
As Figuras 6 e 7 mostram o órgão de Corti que, como já referido, aloja as células ciliadas, assentando sobre a membrana basilar e seguindo a estrutura em espiral da mesma. Como pudemos já verificar, um som normal pode ser obtido como a soma de sons elementares com freqüências diversas. Devido às características muito particulares da cóclea, cada uma dessas freqüências excita uma determinada zona da membrana basilar, estimulando assim apenas as células ciliadas que aí se encontram. Esta particularidade, explorada de forma mais aprofundada no tópico seguinte, constitui a razão da nossa capacidade em diferenciar sons de tonalidades (ou freqüências) diferentes.
|
||||||||||||
| autores: Stephan Blatrix, Rémy Pujol | ||||||||||||
| Figura 6- Desenho de um pormenor de um corte transversal da cóclea. |
Podemos verificar, analisando a Figura 7, que existem dois tipos de células ciliadas: as células ciliadas interiores (1) e as exteriores (2). É também possível verificar que a membrana tectória (6), imersa em endolinfa, cobre os dois tipos de células. Há três filas de aproximadamente 12000 células ciliadas exteriores. Embora sejam em muito maior número do que as células interiores, recebem apenas aproximadamente 5% das inervações das fibras do nervo auditivo. Estas células contêm uma espécie de filamentos musculosos que contraem quando estimulados e “afinam” a resposta da membrana basilar. Por causa deste efeito, as células ciliadas exteriores saudáveis soarão no seguimento de um estímulo sonoro. Ou seja, elas próprias produzem um pequeno som depois de serem estimuladas. Esta é também a razão que está na origem de certas perturbações, causando os conhecidos zumbidos (tinnitus).
As células ciliadas internas distribuem-se ao longo de uma fila com aproximadamente 3500 células. Estas células recebem aproximadamente 95% das enervações das fibras do nervo auditivo, sendo as principais responsáveis pela produção da sensação de audição. Quando danificadas provocam perdas auditivas acentuadas e irreversíveis.
|
||||||||||||||
| autores: Stephan Blatrix, Rémy Pujol | ||||||||||||||
| Figura 7- Pormenor esquemático de uma secção transversal do órgão de Corti.
Neste desenho esquemático, os dois tipos de células sensoriais (interna (CCI – 1) e externas |
Nas Figuras 6 e 7 é ainda possível observar a chamada membrana tectória (6), uma membrana delicada, flexível e gelatinosa que cobre as células ciliadas interiores e exteriores. Os cílios (tufos parecidos com cabelo que se estendem à superfície das células ciliadas) das células exteriores estão encaixados na membrana tectorial. Nas células interiores, os cílios podem ou não estar encaixados na membrana tectorial. Quando um estímulo sonoro provoca oscilações na membrana basilar, a membrana tectória move-se, estimulando assim os cílios.
As Células Ciliadas
As células cocleares são chamadas de células ciliadas, porque são caracterizadas por possuírem uma placa cuticular, onde existe um tufo de cílios, banhados pela endolinfa circundante. O corpo da célula propriamente dito localiza-se no compartimento da perilinfa (veja figura 7). Esquematicamente, os dois tipos de células, (células ciliadas internas (CCI) e células ciliadas externas (CCE)), diferem pelo seu formato e o padrão de seus cílios.
| CCI | 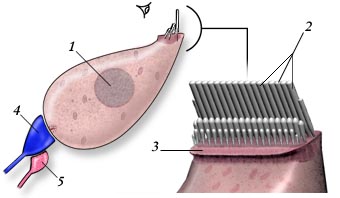 |
|
||||||||
| CCE | 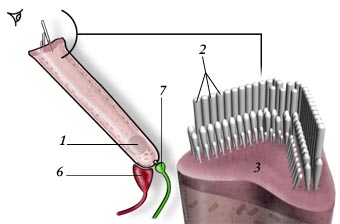 |
|||||||||
| Figura 8 – Detalhe das Células Ciliadas | ||||||||||
Na cóclea humana, existem 3.500 CCI e cerca de 12.000 CCE. Este número é ridiculamente pequeno, quando comparado aos milhões de foto-receptores na retina ou quimio-receptores no nariz! Além disso, as células ciliadas compartilham com os neurônios uma incapacidade de se proliferar, uma vez que se tenham diferenciado. Isto significa que o número final de células ciliadas é alcançado muito precocemente no desenvolvimento (por volta da décima semana da gestação fetal); deste estágio em diante, a nossa cóclea pode apenas perder células ciliadas.
Os cílios das células ciliadas são os responsáveis pela mecano-transdução, ou seja, a tradução da vibração sonora em mensagens nervosas (bio-elétricas), as quais podem ser interpretadas pelo sistema nervoso central.
Arranjo das cílios (ligações)
 |
Os cílios (cerca de uma centena) são geralmente arranjados em três fileiras, seguindo graduações de comprimento. Em adição às pequenas ligações superiores (mostradas aqui em vermelho), as quais estão envolvidas no processo de mecano-transdução, os cílios estão conectados através de ligações transversais (laterais), tanto na mesma fileira (mostrados em laranja) quanto de uma fileira para outra (mostrados em azul) |
| Figura 9 – Padrão do arranjo dos cílios das células ciliadas na cóclea de um mamífero adulto. | |
Ligações Ciliares
| A ligação superior (seta vermelha) e uma ligação lateral (seta azul) entre o cílio médio e o mais alto são claramente visíveis. Em ambas as extremidades da ligação superior, é observada uma condensação da membrana. Estas estruturas estão envolvidas no processo de mecano-transdução. Note os filamentos de actina arranjados de forma regular no interior do cílio (escala: 300 nm). | 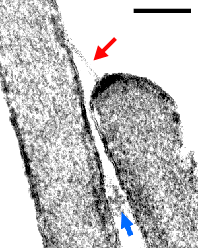 |
| Figura 10 – Ligações superiores (seta vermelha) e ligações entre dois cílios (seta azul) (imagem por microscopia eletrônica) |
|
Processo de Mecano-Transdução
| A despolarização da célula ciliada é baseada na abertura mecânica dos canais catiônicos, provavelmente localizados no topo dos cílios. As ligações superiores permitem uma abertura rápida, de forma sincronizada, para todos os cílios, quando estes são curvados na direção da stria vascularis (veja figura 6). Devido à alta concentração de potássio (K+) na endolinfa, este penetra na célula e despolariza a sua membrana. | |
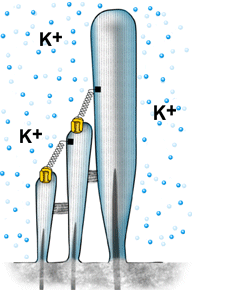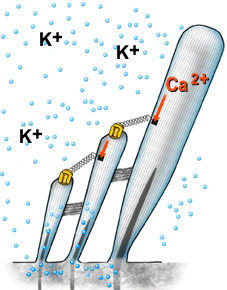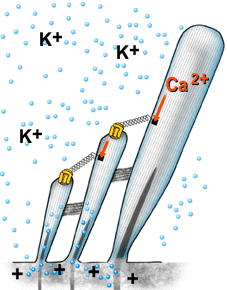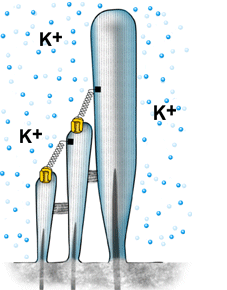 |
Animação Quando os cílios são curvados na direção da stria vascularis, K+ penetra no canal e despolariza a célula. O fechamento do canal ocorre antes do retorno do cílio à sua posição inicial. Este mecanismo é ativado pelo Ca2+ (sua concentração interna aumenta quando os canais são abertos), o qual aciona uma proteína motora (miosina), a qual puxa para baixo as ligações superiores. Este mecanismo reduz a constante temporal da abertura do canal, permitindo desta forma que os ciclos de mecano-transdução ocorram em uma rápida sucessão, ou seja, em altas freqüências. |
| Figura 11 – Animação do processo de mecano-transdução | |
Diagrama de Funcionamento do Órgão de Corti
| OBS – caso a animação não inicie automaticamente, clique com o botão direito e selecione “Reproduzir” ou “Play” | |
O funcionamento básico do órgão de Corti pode ser resumido esquematicamente em 5 estágios:
|
|
| Figura 12 – Animação do funcionamento do Órgão de Corti | |
Fontes:
http://telecom.inescn.pt/research/audio/cienciaviva/constituicao_aaudicao.html
http://www.cochlea.org/po/
